

На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Строение и функции растительной клетки
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
Строение растительной клетки
Строение и функции органоидов растительной клетки
| Органоид | Рисунок | Описание | Функция | Особенности |
Клеточная стенка или плазматическая мембрана | Бесцветная, прозрачная и очень прочная | Пропускает в клетку и выпускает из клетки вещества. | Клеточная мембрана полупроницаемая |
|
Цитоплазма | Густое тягучее вещество | В ней располагаются все другие части клетки | Находится в постоянном движении |
|
Ядро (важная часть клетки) | Округлое или овальное | Обеспечивает передачу наследственных свойств дочерним клеткам при делении | Центральная часть клетки |
|
Сферической или неправильной формы | Принимает участие в синтезе белка | |||
 | Резервуар, отделённый от цитоплазмы мембраной. Содержит клеточный сок | Накапливаются запасные питательные вещества и продукты жизнедеятельности ненужные клетке. | По мере роста клетки мелкие вакуоли сливаются в одну большую (центральную) вакуоль |
|
Пластиды | Хлоропласты | Используют световую энергию солнца и создают органические из неорганических | Форма дисков, отграниченных от цитоплазмы двойной мембраной |
|
Хромопласты | Образуются в результате накопления каротиноидов | Жёлтые, оранжевые или бурые |
||
 | Лейкопласты | Бесцветные пластиды | ||
Ядерная оболочка | Состоит из двух мембран (наружная и внутренняя) с порами | Отграничивает ядро от цитоплазмы | Даёт возможность осуществляться обмену между ядром и цитоплазмой |
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.
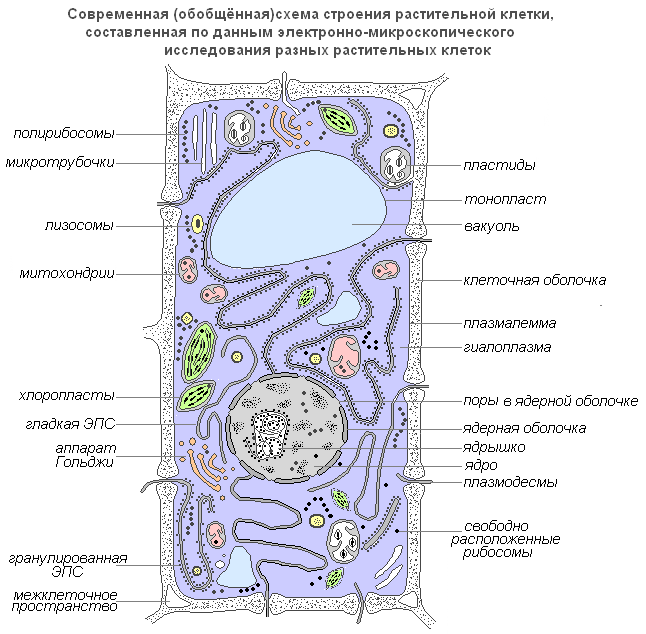
Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.
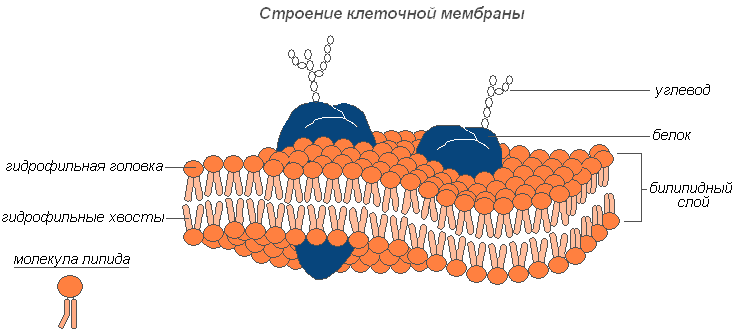
Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Цитоплазматические образования – органеллы
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.

Строение ядра
Ядрышко
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.

Аппарат Гольджи
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Лизосомы
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).

Строение лизосомы
Микротрубочки
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.

Строение микротрубочки
Вакуоль
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.

Строение вакуоли
Пластиды
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.

Строение хлоропласта
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.

Строение лейкопласта
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.

Строение хромопласта
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.

Строение митохондрии
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.

Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.
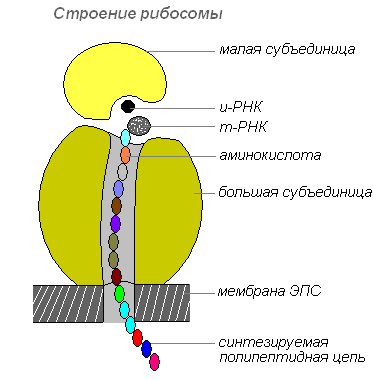
Строение рибосомы
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.
Пластиды – это мембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов (высшие растения, низшие водоросли, некоторые одноклеточные организмы).
Они имеют оболочку, образованную двумя мембранами: наружной и внутренней. Внутренняя мембрана вдается в полость хлоропласта немногочисленными выростами. Мембранная оболочка отграничивает от гиалоплазмы клетки матрикс хлоропласта, так называемую строму. Как строма, так и выросты внутренней мембраны формируют в полости хлоропласта сложную систему мембранных поверхностей, отграничивающих особые плоские мешки, называемые тилакоидами, или ламеллами. Группы дисковидных тилакоидов связаны друг с другом таким образом, что их полости оказываются непрерывными. Эти тилакоиды образуют стопки (наподобие стопки монет), или граны. Тилакоиды стромы объединяют граны между собой. В мембранах тилакоидов сосредоточен главнейший пигмент зеленых растений - хлорофилл и вспомогательные пигменты - каротиноиды. Внутренняя структура хромопластов и лейкопластов проще. Граны в них отсутствуют.
Пластиды высших растений бывают трех видов: хлоропласты, хромопласты и лейкопласты. Они различаются по окраске, строению и выполняемым функциям.
Хлоропласты зеленого цвета, из-за пигмента зеленого цвета – хлорофилла, играющего важную роль в процессах фотосинтеза. Кроме хлорофиллов хлоропласты содержат и каротиноиды, но их гораздо меньше, поэтому они малозаметны на фоне хлорофилла. Больше всего хлоропластов находится в клетках листьев, молодых побегов и незрелых плодов.
Хлоропласты имеют постоянную линзовидную форму. Внутренняя мембрана хлоропластов образует плоские мешочки - тилакоиды (ламеллы). Тилакоиды могут собираться в стопочки – граны. Хлорофилл сосредоточен, главным образом, в тилакоидах гран. В гранах осуществляется световая часть фотосинтеза. Таковая слоистая структура обеспечивает максимальную площадь поверхности мембран и упрощает захват и перенос энергии в процессе фотосинтеза.
Темновая фаза протекает в матриксе хлоропластов, где находятся ферменты, участвующие в этом процессе. Также как и митохондрии, пластиды полуавтономные структуры, имеют свои рибосомы и ДНК.
В хлоропластах может откладываться первичных крахмал, если по каким либо причинам продукты фотосинтеза моносахара длительное время не затребованы клеткой и не удаляются из него.
Характерны для растительных клеток органеллы фотосинтеза, способные создавать из неорганических веществ (CO2 и H2O) при наличии световой энергии и пигмента хлорофилла органические вещества – углеводы и свободный кислород. Синтез собственных белков. Могут образовываться из пластид или лейкопластов, а осенью перейти в хлоропласты (красные и оранжевые плоды, красные и желтые листья).
Хромопласты сосредоточены в цитоплазме клеток плодов, листьев растений и придают им подобающую окраску. Хромопласты образуются из лейкопластов, либо хлоропластов в итоге скопления пигментов каротиноидов. Хромопласты имеют окраску от желтого до оранжевого из-за того, что накапливают пигменты каротиноиды. Также как и у лейкопластов у хлоропластов внутренняя мембрана не развита. Форма хромопластов может быть самой разнообразной: от сферической (каротиноиды откладываются в виде жировых капель) до многогранной (пигменты откладываются в виде кристаллов). Считается, что хромопласты – это конечный этап развития пластид. Они встречаются в клетках созревших плодов, стареющих листьев, т.е. в таких органах, где снижена активность жизнедеятельности органа. Однако они обнаруживаются и в вполне функционирующих частях, таких как корнеплоды моркови. Функция хромопластов пока не установлена.
Характерны для растительных клеток. Придают лепесткам цветков окраску, привлекательную для насекомых-опылителей. В осенних листьях и зрелых плодах отделяющихся от растений, содержатся кристаллические каротиноиды?– конечные продукты обмена.
Лейкопласты - бесцветные пластиды, располагающиеся в неокрашенных частях растений: в стеблях, корнях, луковицах и др. Основная функция лейкопластов – накопление запасных веществ, поэтому у них слабо развита внутренняя мембрана, она почти не образует тилакоидов. Чаще всего в лейкопластах накапливаются зерна вторичного крахмала, такие пластиды называются амилопластами. В них могут также откладываться масла (элайопласты) и простые белки (протеинопласты). Форма лейкопластов непостоянна и зависит от вида накапливаемых веществ. Лейкопласты могут образовываться из хлоропластов при значительном снижении интенсивности освещения.
Характерны для растительных клеток. Служат местом отложения запасных питательных веществ, главным образом крахмальных зерен.
Хлоропласты
– это зеленые пластиды высших растений, содержащие хлорофилл – фотосинтезирующий пигмент. Представляют собой тельца округлой формы размерами от 4 до 10 мкм. Химический состав хлоропласта: примерно 50% белка, 35% жиров, 7% пигментов, малое количество ДНК и РНК. У представителей разных групп растений комплекс пигментов, определяющих окраску и принимающих участие в фотосинтезе, отличается. Это подтипы хлорофилла и каротиноиды (ксантофилл и каротин). При рассматривании под световым микроскопом видна зернистая структура пластид – это граны. Под электронным микроскопом наблюдаются небольшие прозрачные уплощенные мешочки (цистерны, или граны), образованные белково-липидной мембраной и располагающиеся в непосредственно в строме. Причем некоторые из них сгруппированы в пачки, похожие на столбики монет (тилакоиды гран), другие, более крупные находятся между тилакоидами. Благодаря такому строению, увеличивается активная синтезирующая поверхность липидно-белково-пигментного комплекса гран, в котором на свету происходит фотосинтез.
Хромопласты
Лейкопласты
представляют собой бесцветные пластиды, основная функция которых обычно запасающая. Размеры этих органелл относительно небольшие. Они округлой либо слегка продолговатой формы, характерны для всех живых клеток растений. В лейкопластах осуществляется синтез из простых соединений более сложных – крахмала, жиров, белков, которые сохраняются про запас в клубнях, корнях, семенах, плодах. Под электронным микроскопом заметно, что каждый лейкопласт покрыт двухслойной мембраной, в строме есть только один или небольшое число выростов мембраны, основное пространство заполнено органическими веществами. В зависимости от того, какие вещества накапливаются в строме, лейкопласты делят на амилопласты, протеинопласты и элеопласты.
74. Каково строение ядра, роль в клетке? Какие структуры ядра обуславливают его функции? Что такое хроматин?
Ядро – основной компонент клетки, несущей генетическую информации Ядро – располагается в центре. Форма различная, но всегда круглая или овальная. Размеры различны. Содержимое ядра – жидкая консистенция. Различают оболочку, хроматин, кариолимфу (ядерный сок), ядрышко. Ядерная оболочка состоит из 2 мембран, разделённых перенуклеарным пространством. Оболочка снабжена порами, через которые происходит обмен крупными молекулами различных веществ. Оно может находиться в 2 состояниях: покоя – интерфазы и деления – митоза или мейоза.
Ядро осуществляет две группы общих функций: одну, связанную собственно с хранением генетической информации, другую — с ее реализацией, с обеспечением синтеза белка.
В первую группу входят процессы, связанные с поддержанием наследственной информации в виде неизменной структуры ДНК. Эти процессы связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекулы ДНК (разрыв одной из цепей ДНК, часть радиационных повреждений), что сохраняет строение молекул ДНК практически неизменным в ряду поколений клеток или организмов. Далее, в ядре происходит воспроизведение или редупликация молекул ДНК, что дает возможность двум клеткам получить совершенно одинаковые и в качественном и в количественном смысле объемы генетической информации. В ядрах происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток.
Другой группой клеточных процессов, обеспечивающихся активностью ядра, является создание собственно аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК и рибосомных РНК. В ядре эукариотов происходит также образование субъедениц рибосом путем комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро.
Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится. Поэтому выпадение лил нарушение любой из перечисленных выше функций губительно для клетки в целом. Так нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не будет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что также губительно для клеток. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток. Выпадение в результате поражения ядра или в случае нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке или к грубым его нарушениям.
Хроматин
(греч. χρώματα - цвета, краски) - это вещество хромосом - комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК.
75. Каково строение и типы хромосом? Что такое кариотип, аутосомы, гетеросомы, диплоидный и гаплоидный наборы хромосом?
Хромосомы – органоиды клеточного ядра, совокупность которых определяет основные наследственные свойства клеток и организмов. Полный набор хромосом в клетке, характерный для данного организма, называется кариотипом. В любой клетке тела большинства животных и растений каждая хромосома представлена дважды: одна из них получена от отца, другая – от матери при слиянии ядер половых клеток в процессе оплодотворения. Такие хромосомы называются гомологичными, набор гомологичных хромосом – диплоидным. В хромосомном наборе клеток раздельнополых организмов присутствует пара (или несколько пар) половых хромосом, как правило, различающихся у разных полов по морфологическим признакам; остальные хромосомы называются аутосомами. У млекопитающих в половых хромосомах локализованы гены, определяющие пол организма.
Значение хромосом как клеточных органоидов, ответственных за хранение, воспроизведение и реализацию наследственной информации, определяется свойствами биополимеров, входящих в их состав.
Аутосомами
у живых организмов с хромосомным определением пола называют парные хромосомы, одинаковые у мужских и женских организмов. Иными словами, кроме половых хромосом, все остальные хромосомы у раздельнополых организмов будут являться аутосомами.
Аутосомы обозначают порядковыми номерами. Так, у человека в диплоидном наборе имеется 46 хромосом, из них - 44 аутосомы (22 пары, обозначаемые номерами с 1-го по 22-й) и одна пара половых хромосом (XX у женщин и XY у мужчин).
Гаплоидный набор хромосом Начнем с гаплоидного. Он представляет собой скопление совершенно разных хромосом, т.е. в организме-гаплоиде есть несколько этих нуклеопротеидных структур, непохожих друг на друга (фото). Гаплоидный набор хромосом характерен для растений, водорослей и грибов. Диплоидный набор хромосом Этот набор является таким собранием хромосом, при котором у каждой из них есть двойник, т.е. эти нуклепротеидные структуры расположены попарно (фото). Диплоидный набор хромосом характерен для всех животных, в том числе и человека. Кстати, о последнем. У здорового человека их 46, т.е. 23 пары. Однако его пол определяют всего две, называемые половыми, — Х и Y- Читайте подробнее на SYL.ru:
76. Дайте определение клеточного цикла, охарактеризуйте его фазы. Какие функции жизни обеспечиваются делением клеток?
Клеточный цикл - это период существования клетки от момента её образования путем деления материнской клетки до собственного деления или гибели.
Клеточный цикл эукариот состоит из двух периодов:
1Период клеточного роста, называемый «интерфаза», во время которого идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
2Периода клеточного деления, называемый «фаза М» (от слова mitosis - митоз).
Деление клетки. Рост организма осуществляется за счет деления его клеток. Способность к делению — важнейшее свойство клеточной жизнедеятельности. Делясь, клетка удваивает все свои структурные компоненты, и в результате возникают две новые клетки. Наиболее распространенным способом деления клетки является митоз — непрямое деление клетки.
Предыдущая24252627282930313233343536373839Следующая
Пластиды
Пластиды являются основными цитоплазматическими органеллами клеток автотрофных растений. Название происходит от греческого слова «plastos», что в переводе означает «вылепленный».
Главная функция пластид – синтез органических веществ, благодаря наличию собственных ДНК и РНК и структур белкового синтеза. В пластидах также содержатся пигменты, обусловливающие их цвет. Все виды данных органелл имеют сложное внутреннее строение. Снаружи пластиду покрывают две элементарные мембраны, имеется система внутренних мембран, погруженных в строму или матрикс.
Классификация пластид по окраске и выполняемой функции подразумевает деление этих органоидов на три типа: хлоропласты, лейкопласты и хромопласты. Пластиды водорослей именуются хроматофорами.
Хлоропласты – это зеленые пластиды высших растений, содержащие хлорофилл – фотосинтезирующий пигмент. Представляют собой тельца округлой формы размерами от 4 до 10 мкм. Химический состав хлоропласта: примерно 50% белка, 35% жиров, 7% пигментов, малое количество ДНК и РНК. У представителей разных групп растений комплекс пигментов, определяющих окраску и принимающих участие в фотосинтезе, отличается. Это подтипы хлорофилла и каротиноиды (ксантофилл и каротин). При рассматривании под световым микроскопом видна зернистая структура пластид – это граны. Под электронным микроскопом наблюдаются небольшие прозрачные уплощенные мешочки (цистерны, или граны), образованные белково-липидной мембраной и располагающиеся в непосредственно в строме.
Причем некоторые из них сгруппированы в пачки, похожие на столбики монет (тилакоиды гран), другие, более крупные находятся между тилакоидами. Благодаря такому строению, увеличивается активная синтезирующая поверхность липидно-белково-пигментного комплекса гран, в котором на свету происходит фотосинтез.
Хромопласты – пластиды, окраска которых бывает желтого, оранжевого или красного цвета, что обусловлено накоплением в них каротиноидов. Благодаря наличию хромопластов, характерную окраску имеют осенние листья, лепестки цветов, созревшие плоды (помидоры, яблоки). Данные органоиды могут быть различной формы – округлой, многоугольной, иногда игольчатой.
Лейкопласты представляют собой бесцветные пластиды, основная функция которых обычно запасающая. Размеры этих органелл относительно небольшие.
Они округлой либо слегка продолговатой формы, характерны для всех живых клеток растений. В лейкопластах осуществляется синтез из простых соединений более сложных – крахмала, жиров, белков, которые сохраняются про запас в клубнях, корнях, семенах, плодах. Под электронным микроскопом заметно, что каждый лейкопласт покрыт двухслойной мембраной, в строме есть только один или небольшое число выростов мембраны, основное пространство заполнено органическими веществами. В зависимости от того, какие вещества накапливаются в строме, лейкопласты делят на амилопласты, протеинопласты и элеопласты.
Все виды пластид имеют общее происхождение и способны переходить из одного вида в другой. Так, превращение лейкопластов в хлоропласты наблюдается при позеленении картофельных клубней на свету, а в осенний период в хлоропластах зеленых листьев разрушается хлорофилл, и они трансформируются в хромопласты, что проявляется пожелтением листьев. В каждой определенной клетке растения может быть только один вид пластид.
Пластиды - это органоиды клеток растений и некоторых фотосинтезирующих простейших. У животных и грибов пластид нет.
Пластиды делятся на несколько типов. Наиболее важный и известный - хлоропласт, содержащий зеленый пигмент хлорофилл, который обеспечивает процесс фотосинтеза.
Другими видами пластид являются разноцветные хромопласты и бесцветные лейкопласты. Также выделяют амилопласты, липидопласты, протеинопласты, которые часто считают разновидностями лейкопластов.
Все виды пластид связаны между собой общим происхождением или возможным взаимопревращением. Пластиды развиваются из пропластид – более мелких органоидов меристематических клеток.
Строение пластид
Большинство пластид относится к двумембранным органоидам, у них есть внешняя и внутренняя мембраны.
Однако встречаются организмы, чьи пластиды имеют четыре мембраны, что связано с особенностями их происхождения.
Во многих пластидах, особенно в хлоропластах, хорошо развита внутренняя мембранная система, формирующая такие структуры как тилакоиды, граны (стопки тилакоидов), ламелы – удлиненные тилакоиды, соединяющие соседние граны. Внутренне содержимое пластид обычно называют стромой. В ней помимо прочего находятся крахмальные зерна.
Считается, что в процессе эволюции пластиды появились аналогично митохондриям - путем внедрения в клетку-хозяина другой прокариотической клетки, способной в данном случае к фотосинтезу. Поэтому пластиды считают полуавтономными органеллами. Они могут делиться независимо от делений клетки, у них есть собственная ДНК, РНК, рибосомы прокариотического типа, т. е. собственный белоксинтезирующий аппарат. Это не значит, что в пластиды не поступают белки и РНК из цитоплазмы. Часть генов, управляющей их функционированием, находится как раз в ядре.
Функции пластид
Функции пластид зависят от их типа. Хлоропласты выполняют фотосинтезирующую функцию. В лейкопластах накапливаются запасные питательные вещества: крахмал в амилопластах, жиры в элайопластах (липидопластах), белки в протеинопластах.
Хромопласты, за счет содержащихся в них пигментов-каротиноидов, окрашивают различные части растений – цветки, плоды, корнеплоды, осенние листья и др. Яркий окрас часто служит своеобразным сигналом для животных-опылителей и распространителей плодов и семян.
В дегенерирующих зеленых частях растений хлоропласты превращаются в хромопласты. Пигмент хлорофилл разрушается, поэтому остальные пигменты, несмотря на малое количество, становятся в пластидах заметными и окрашивают туже листву в желто-красные оттенки.
Пластиды - это органоиды растительных клеток. Одним из видов пластид являются фотосинтезирующие хлоропласты. Другие распространенные разновидности - хромопласты и лейкопласты. Все их объединяет единство происхождения и общий план строения. Различает - преобладание определенных пигментов и выполняемые функции.
Пластиды развиваются из пропластид, которые присутствуют в клетках образовательной ткани и существенно меньше по размеру, чем зрелый органоид. Кроме того, пластиды способны к делению надвое перетяжкой, что подобно делению бактерий.
В строении пластид выделяют внешнюю и внутреннюю мембраны, внутреннее содержимое - строму, внутреннюю мембранную систему, которая особенно развита в хлоропластах, где формирует тилакоиды, граны и ламелы.
В строме содержится ДНК, рибосомы, различные типы РНК. Таким образом, как и митохондрии, пластиды способны к самостоятельному синтезу части необходимых белковых молекул. Считается, что в процессе эволюции пластиды и митохондрии появились в результате симбиоза разных прокариотических организмов, один из которых стал клеткой-хозяином, а другие - ее органеллами.
Функции пластид зависят от их вида:
- хлоропласты → фотосинтез,
- хромопласты → окраска частей растения,
- лейкопласты → запас питательных веществ.
Растительные клетки содержат преимущественно один из видов пластид. В хлоропластах преобладает пигмент хлорофилл, поэтому содержащие их клетки зеленые. В хромопластах содержатся пигменты каротиноиды, которые придают цвет от желтого, через оранжевый к красному.
Лейкопласты бесцветны.
Окраска хромопластами цветков и плодов растения в яркие цвета привлекает насекомых-опылителей и животных-распространителей семян. В осенних листьях происходит разрушение хлорофилла, в результате цвет определяется каротиноидами. Из-за этого листва приобретает соответствующую окраску. При этом хлоропласты превращаются в хромопласты, которые часто рассматривают как конечную стадию развития пластид.
Лейкопласты при освещении способны превращаться в хлоропласты. Это можно наблюдать у клубней картофеля, когда на свету они начинают зеленеть.
Выделяют несколько видов лейкопластов в зависимости от типа накапливаемых в них веществ:
- протеинопласты → белки,
- элайопласты , или липидопласты, → жиры,
- амилопласты → углеводы, обычно в виде крахмала.
Лекция № 6.
Количество часов: 2
МИТОХОНДРИИ И ПЛАСТИДЫ
1.
2. Пластиды, строение, разновидности, функции
3.
Митохондрии и пластиды – двухмембранные органоиды эукариотических клеток. Митохондрии встречаются во всех клетках животных и растений. Пластиды характерны для клеток растений, осуществляющих фотосинтетические процессы. Эти органоиды имеют сходный план строения и некоторые общие свойства. Однако по основным метаболическим процессам они существенно отличаются друг от друга.
1. Митохондрии, строение, функциональное значение
Общая характеристика митохондрий. Митохондрии (греч. “митос” - нить, “хондрион” - зерно, гранула) – округлые, овальные или палочковидные двухмембранные органоиды диаметром около 0,2-1 мкм и длиной до 7-10 мкм. Эти органоиды можно обнаружить с помощью световой микроскопии, поскольку они обладают достаточной величиной и высокой плотностью. Особенности внутреннего строения их можно изучить только с помощью электронного микроскопа. Митохондрии были открыты в 1894 г. Р. Альтманом, который дал им название «биобласты». Термин "митохондрия" был введен К. Бенда в 1897 г. Митохондрии имеются практически во всех эукариотических клетках. У анаэробных организмов (кишечные амебы и др.) митохондрии отсутствуют. Число митохондрий в клетке колеблется от 1 до 100 тыс. и зависит от типа, функциональной активности и возраста клетки. Так в растительных клетках митохондрий меньше, чем в животных; а в молодых клетках больше, чем в старых. Жизненный цикл митохондрий составляет несколько дней. В клетке митохондрии обычно скапливаются вблизи участков цитоплазмы, где возникает потребность в АТФ. Например, в сердечной мышце митохондрии находятся вблизи миофибрилл, а в спермиях образуют спиральный футляр вокруг оси жгутика.
Ультрамикроскопическое строение митохондрий. Митохондрии ограничены двумя мембранами, каждая из которых имеет толщину около 7 нм. Внешнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внешняя мембрана гладкая, а внутренняя образует складки – кристы (лат. “криста” – гребень, вырост), увеличивающие ее поверхность. Число крист неодинаково в митохондриях разных клеток. Их может быть от нескольких десятков до нескольких сотен. Особенно много крист в митохондриях активно функционирующих клеток, например мышечных. В кристах располагаются цепи переноса электронов и сопряженного с ним фосфорилирования АДФ (окислительное фосфорилирование). Внутреннее пространство митохондрий заполнено гомогенным веществом, называемым матриксом. Митохондриальные кристы обычно полностью не перегораживают полость митохондрии. Поэтому матрикс на всем протяжении является непрерывным. В матриксе содержатся кольцевые молекулы ДНК, митохондриальные рибосомы, встречаются отложения солей кальция и магния. На митохондриальной ДНК происходит синтез молекул РНК различных типов, рибосомы участвуют в синтезе ряда митохондриальных белков. Малые размеры ДНК митохондрий не позволяют кодировать синтез всех митохондриальных белков. Поэтому синтез большинства белков митохондрий находится под ядерным контролем и осуществляется в цитоплазме клетки. Без этих белков рост и функционирование митохондрий невозможно. Митохондриальная ДНК кодирует структурные белки, ответственные за правильную интеграцию в митохондриальных мембранах отдельных функциональных компонентов.
Размножение митохондрий. Митохондрии размножаются путем деления перетяжкой или фрагментацией крупных митохондрий на более мелкие. Образовавшиеся таким путем митохондрии могут расти и снова делиться.
Функции митохондрий. Основная функция митохондрий заключается в синтезе АТФ. Этот процесс происходит в результате окисления органических субстратов и фосфорилирования АДФ. Первый этап этого процесса происходит в цитоплазме в анаэробных условиях. Поскольку основным субстратом является глюкоза, то процесс носит название гликолиза. На данном этапе субстрат подвергается ферментативному расщеплению до пировиноградной кислоты с одновременным синтезом небольшого количества АТФ. Второй этап происходит в митохондриях и требует присутствия кислорода. На этом этапе происходит дальнейшее окисление пировиноградной кислоты с выделением СО 2 и переносом электронов на акцепторы. Эти реакции осуществляются с помощью ряда ферментов цикла трикарбоновых кислот, которые локализованы в матриксе митохондрии. Освободившиеся в процессе окисления в цикле Кребса электроны переносятся в дыхательную цепь (цепь переноса электронов). В дыхательной цепи они соединяются с молекулярным кислородом, образуя молекулы воды. В результате этого небольшими порциями выделяется энергия, которая запасается в виде АТФ. Полное окисление одной молекулы глюкозы с образованием диоксида углерода и воды обеспечивает энергией перезарядку 38 молекул АТФ (2 молекулы в цитоплазме и 36 в митохондриях).
Аналоги митохондрий у бактерий. У бактерий митохондрий нет. Вместо них у них имеются цепи переноса электронов, локализованные в мембране клетки.
2. Пластиды, строение, разновидности, функции. Проблема происхождения пластид
Пластиды (от. греч. plastides – создающие, образующие) – это двухмембранные органоиды, характерные для фотосинтезирующих эукариотных организмов. Различают три основных типа пластид: хлоропласты, хромопласты и лейкопласты. Совокупность пластид в клетке называют пластидомом. Пластиды связаны между собой единым происхождением в онтогенезе от пропластид меристематических клеток. Каждый их этих типов при определенных условиях может переходить один в другой. Как и митохондрии, пластиды содержат собственные молекулы ДНК. Поэтому они также способны размножаться независимо от деления клетки.
Хлоропласты (от греч. « chloros » – зеленый, « plastos » - вылепленный) – это пластиды, в которых осуществляется фотосинтез.
Общая характеристика хлоропластов. Хлоропласты представляют собой органоиды зеленого цвета длиной 5-10 мкм и шириной 2-4 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм. У высших растений хлоропласты имеют двояковыпуклую или эллипсоидную форму. Количество хлоропластов в клетке может варьировать от одного (некоторые зеленые водоросли) до тысячи (махорка). В клетке высших растений в среднем находится 15-50 хлоропластов. Обычно хлоропласты равномерно распределены по цитоплазме клетки, но иногда они группируются около ядра или клеточной оболочки. По-видимому, это зависит от внешних воздействий (интенсивность освещения).
Ультрамикроскопическое строение хлоропластов. От цитоплазмы хлоропласты отделены двумя мембранами, каждая из которых имеет толщину около 7 нм. Между мембранами находится межмембранное пространство диаметром около 20-30 нм. Наружная мембрана гладкая, внутренняя имеет складчатую структуру. Между складками располагаются тилакоиды , имеющие вид дисков. Тилакоиды образуют стопки наподобие столбика монет, называемые гранами. М ежду собой граны соединены другими тилакоидами (ламелы, фреты ). Число тилакоидов в одной гране варьирует от нескольких штук до 50 и более. В свою очередь в хлоропласте высших растений находится около 50 гран (40-60), расположенных в шахматном порядке. Такое расположение обеспечивает максимальную освещенность каждой граны. В центре граны находится хлорофилл, окруженный слоем белка; затем располагается слой липоидов, снова белок и хлорофилл. Хлорофилл имеет сложное химическое строение и существует в нескольких модификациях (a , b , c , d ). У высших растений и водорослей в качестве основного пигмента содержится х лорофилл а с формулой С 55 Н 72 О 5 N 4 М g . В качестве дополнительных содержатся хлорофилл b (высшие растения, зеленые водоросли), хлорофилл с (бурые и диатомовые водоросли), хлорофилл d (красные водоросли). Образование хлорофилла происходит только при наличии света и железа, играющего роль катализатора. Матрикс хлоропласта представляет собой бесцветное гомогенное вещество, заполняющее пространство между тилакоидами. В матриксе находятся ферменты "темновой фазы" фотосинтеза, ДНК, РНК, рибосомы. Кроме этого, в матриксе происходит первичное отложение крахмала в виде крахмальных зерен.
Свойства хлоропластов:
· полуавтономность (имеют собственный белоксинтезирующий аппарат, однако большая часть генетической информации находится в ядре);
· способность к самостоятельному движению (уходят от прямых солнечных лучей);
· способность к самостоятельному размножению.
Размножение хлоропластов. Хлоропласты развиваются из пропластид, которые способны реплицироваться путем деления. У высших растений также встречается деление зрелых хлоропластов, но крайне редко. При старении листьев и стеблей, созревании плодов хлоропласты утрачивают зеленую окраску, превращаясь в хромопласты.
Функции хлоропластов. Основная функция хлоропластов – фотосинтез. Кроме фотосинтеза хлоропласты осуществляют синтез АТФ из АДФ (фосфорилирование), синтез липидов, крахмала, белков. В хлоропластах также синтезируются ферменты, обеспечивающие световую фазу фотосинтеза.
Хромопласты (от греч. chromatos – цвет, краска и « plastos » – вылепленный) – это окрашенные пластиды. Цвет их обусловлен наличием следующих пигментов: каротина (оранжево-желтый), ликопина (красный) и ксантофилла (желтый). Хромопластов особенно много в клетках лепестков цветков и оболочек плодов. Больше всего хромопластов в плодах и увядающих цветках и листьях. Хромопласты могут развиваться из хлоропластов, которые при этом теряют хлорофилл и накапливают каротиноиды. Это происходит при созревании многих фруктов: налившись спелым соком, они желтеют, розовеют или краснеют. Основная функция хромопластов заключается в обеспечении окраски цветов, плодов, семян.
В отличие от лейкопластов и особенно хлоропластов внутренняя мембрана хлоропластов не образует тилакоидов (или образует одиночные). Хромопласты – это конечный итог развития пластид (в хромопласты превращаются хлоропласты и пластиды).
Лейкопласты (от греч. leucos – белый, plastos – вылепленный, созданный) . Это бесцветные пластиды округлой, яйцевидной, веретенообразной формы. Находятся в подземных частях растений, семенах, эпидермисе, сердцевине стебля. Особенно богаты лейкопластами клубни картофеля. Внутренняя оболочка образует немногочисленные тилакоиды. На свету из хлоропластов образуются хлоропласты. Лейкопласты, в которых синтезируется и накапливается вторичный крахмал называют амилопластами , масла – эйлалопластами , белки – протеопластами. Основная функция лейкопластов – это аккумуляция питательных веществ.
3. Проблема происхождения митохондрий и пластид. Относительная автономия
Существует две основные теории происхождения митохондрий и пластид. Это теории прямой филиации и последовательных эндосимбиозов. Согласно теории прямой филиации митохондрии и пластиды образовались путем компартизации самой клетки. Фотосинтезирующие эукариоты произошли от фотосинтезирующих прокариот. У образовавшихся автотрофных эукариотических клеток путем внутриклеточной дифференцировки образовались митохондрии. В результате утраты пластид от автотрофов произошли животные и грибы.
Наиболее обоснованной является теория последовательных эндосимбиозов. Согласно этой теории возникновение эукариотической клетки прошло через несколько этапов симбиоза с другими клетками. На первой стадии клетки типа анаэробных гетеротрофных бактерий включили в себя свободноживущие аэробные бактерии, превратившиеся в митохондрии. Параллельно этому в клетке-хозяине прокариотической генофор формируется в обособленное от цитоплазмы ядро. Таким путем возникла первая эукариотическая клетка, которая была гетеротрофной. Возникшие эукариотические клетки путем повторных симбиозов включили в себя синезеленые водоросли, что привело к появлению в них структур типа хлоропластов. Таким образом, митохондрии уже были у гетеротрофных эукариотических клеток, когда последние в результате симбиоза приобрели пластиды. В дальнейшем в результате естественного отбора митохондрии и хлоропласты утратили часть генетического материала и превратились в структуры с ограниченной автономией.
Доказательства эндосимбиотической теории:
1. Сходство структуры и энергетических процессов у бактерий и митохондрий, с одной стороны, и у синезеленых водорослей и хлоропластов, с другой стороны.
2. Митохондрии и пластиды имеют собственную специфическую систему синтеза белков (ДНК, РНК, рибосомы). Специфичность этой системы заключается в автономности и резком отличии от таковой в клетке.
3. ДНК митохондрий и пластид представляет собой небольшую циклическую или линейную молекулу, которая отличается от ДНК ядра и по своим характеристикам приближается к ДНК прокариотических клеток. Синтез ДНК митохондрий и пластид не зависит от синтеза ядерной ДНК.
4. В митохондриях и хлоропластах имеются и-РНК, т-РНК, р-РНК. Рибосомы и р-РНК этих органоидов резко отличаются от таковых в цитоплазме. В частности рибосомы митохондрий и хлоропластов, в отличие от цитоплазматических рибосом, чувствительны к антибиотику хлорамфениколу, подавляющему синтез белка у прокариотических клеток.
5. Увеличение числа митохондрий происходит путем роста и деления исходных митохондрий. Увеличение числа хлоропластов происходит через изменения пропластид, которые, в свою очередь, размножаются путем деления.
Эта теория хорошо объясняет сохранение у митохондрий и пластид остатков систем репликации и позволяет построить последовательную филогению от прокариот к эукариотам.
Относительная автономия хлоропластов и пластид. В некоторых отношениях митохондрии и хлоропласты ведут себя как автономные организмы. Например, эти структуры образуются только из исходных митохондрий и хлоропластов. Это было продемонстрировано в опытах на растительных клетках, у которых образование хлоропластов подавляли антибиотиком стрептомицином, и на клетках дрожжей, где образование митохондрий подавляли другими препаратами. После таких воздействий клетки уже никогда не восстанавливали отсутствующие органеллы. Причина в том, что митохондрии и хлоропласты содержат определенное количество собственного генетического материала (ДНК), который кодирует часть их структуры. Если эта ДНК утрачивается, что и происходит при подавлении образования органелл, то структура не может быть воссоздана. Оба типа органелл имеют свою собственную белок-синтезирующую систему (рибосомы и транспортные РНК), которая несколько отличается от основной белок-синтезирующей системы клетки; известно, например, что белок-синтезирующая система органелл может быть подавлена с помощью антибиотиков, тогда как на основную систему они не действуют. ДНК органелл ответственна за основную часть внехромосомной, или цитоплазматической, наследственности. Внехромосомная наследственность не подчиняется менделевским законам, так как при делении клетки ДНК органелл передается дочерним клеткам иным путем, нежели хромосомы. Изучение мутаций, которые происходят в ДНК органелл и ДНК хромосом, показало, что ДНК органелл отвечает лишь за малую часть структуры органелл; большинство их белков закодированы в генах, расположенных в хромосомах. Относительная автономия митохондрий и пластид рассматривается как одно из доказательств их симбиотического происхождения.
Пластиды – это мембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов (высшие растения, низшие водоросли, некоторые одноклеточные организмы). Подобно митохондриям, пластиды окружены двумя мембранами, в их матриксе имеется собственная геномная система, функции пластид связаны с энергообеспечением клетки, идущим на нужды фотосинтеза. У высших растений найден целый набор различных пластид (хлоропласт, лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных превращений одного вида пластиды в другой. Основной структурой, которая осуществляет фотосинтетические процессы, является хлоропласт (рис. 226а).
Хлоропласт
Как уже указывалось, строение хлоропласта в принципе напоминает строение митохондрии. Обычно это структуры удлиненной формы с шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм. Количество хлоропластов в клетках разных растений не стандартно. Так, у зеленых водорослей может быть по одному хлоропласту на клетку. Обычно на клетку высших растений приходится в среднем 10-30 хлоропластов. Встречаются клетки с огромным количеством хлоропластов. Например, в гигантских клетках палисадной ткани махорки обнаружено около 1000 хлоропластов.
Хлоропласты представляют собой структуры, ограниченные двумя мембранами – внутренней и внешней. Внешняя мембрана, как и внутренняя, имеет толщину около 7 мкм, они отделены друг от друга межмембранным пространством около 20-30 нм. Внутренняя мембрана хлоропластов отделяет строму пластиды, аналогичную матриксу митохондрий. В строме зрелого хлоропласта высших растений видны два типа внутренних мембран. Это – мембраны, образующие плоские, протяженные ламеллы стромы , и мембраны тилакоидов , плоских дисковидных вакуолей или мешков.
Ламеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешки или же имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
Кроме мембран стромы в хлоропластах обнаруживаются мембранные тилакоиды. Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина межмембранного пространства у них также около 20-30 нм. Такие тилакоиды образуют стопки наподобие столбика монет, называемые гранами (рис. 227). Число тилакоидов на одну грану очень варьирует: от нескольких штук до 50 и более. Размер таких стопок может достигать 0,5 мкм, поэтому граны видны в некоторых объектах в световом микроскопе. Количество гран в хлоропластах высших растений может достигать 40-60. Тилакоиды в гране сближены друг с другом так, что внешние слои их мембран тесно соединяются; в месте соединения мембран тилакоидов образуется плотный слой толщиной около 2 нм. В состав граны кроме замкнутых камер тилакоидов обычно входят и участки ламелл, которые в местах контакта их мембран с мембранами тилакоидов тоже образуют плотные 2-нм слои. Ламеллы стромы, таким образом, как бы связывают между собой отдельные граны хлоропласта. Однако полости камер тилакоидов всезда замкнуты и не переходят в камеры межмембранного пространства ламелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
В матриксе (строме) хлоропластов обнаруживаются молекулы ДНК, рибосомы; там же происходит первичное отложение запасного полисахарида, крахмала, в виде крахмальных зерен.
Функции хлоропластов
Хлоропласты – это структуры, в которых происходят фотосинтетические процессы, приводящие в конечном итоге к связыванию углекислоты, к выделению кислорода и синтезу сахаров.
Характерным для хлоропластов является наличие в них пигментов, хлорофиллов, которые и придают окраску зеленым растениям. При помощи хлорофилла зеленые растения поглощают энергию солнечного света и превращают ее в химическую. Поглощение света с определенной длиной волны приводит к изменению в структуре молекулы хлорофилла, она переходит при этом в возбужденное, активированное состояние. Освобождающаяся энергия активированного хлорофилла через ряд промежуточных этапов передается определенным синтетическим процессам, приводящим к синтезу АТФ и к восстановлению акцептора электронов НАДФ (никотинамидадениндинуклеотид) до НАДФ-Н, которые тратятся на реакции связывания СО 2 и синтез сахаров.
Суммарная реакция фотосинтеза может быть выражена следующим образом:
nСО 2 + nН 2 О свет (СН 2 О) n + nО 2 (I)
хлорофилл
Таким образом, главный итоговый процесс здесь – связывание двуокиси углерода с использование воды для образования различных углеводов и выделение кислорода. Молекулы кислорода, который выделяется в процессе фотосинтеза у растений, образуется за счет гидролиза молекулы воды. Следовательно, процесс фотосинтеза включает в себя процесс гидролиза воды, которая служит одним из источников электронов или атомов водорода. Биохимические исследования показали, что процесс фотосинтеза представляет собой сложную цепь событий, заключающую в себе две фазы: световую и темновую. Первая, протекающая только на свету, связана с поглощением света хлорофиллами и с проведением фотохимической реакции (реакция Хилла). Во второй фазе, которая может идти в темноте, происходит фиксация и восстановление СО 2 , приводящие к синтезу углеводов.
В результате световой фазы происходит фотофосфорилирование, синтез АТФ из АДФ и фосфата с использованием цепи переноса электронов, а также восстановление кофермента НАДФ (никотинамидадениндинуклеотидфосфат) в НАДФ-Н, происходящего при гидролизе и ионизации воды. В этой фазе фотосинтеза энергия солнечного света возбуждает электроны в молекулах хлорофилла, которые расположены в мембранах тилакоидов. Эти возбужденные электроны переносятся по компонентам окислительной цепи в тилакоидной мембране, подобно тому как электроны транспортируются по дыхательной цепи в мембране митохондрий. Энергия, освобождающаяся при таком переносе электронов, используется для перекачивания протонов через тилакоидную мембрану внутрь тилакоидов, что приводит к возрастанию разности потенциалов между стромой и пространством внутри тилакоида. Также как и в мембранах крист митохондрий в мембранах тилакоидов встроены молекулярные комплексы АТФ-синтетазы, которые начинают затем транспортировать протоны обратно в матрикс хлоропласта, или строму, и параллельно этому фосфорилировать АДФ, т.е. синтезировать АТФ (рис. 228, 229).
Таким образом, в результате световой фазы происходит синтез АТФ и восстановление НАДФ, которые затем используются при восстановлении СО 2 , в синтезе углеводов уже в темновой фазе фотосинтеза.
В темновой (независящей от потока фотонов) стадии фотосинтеза за счет восстановленного НАДФ и энергии АТФ происходит связывание атмосферного СО 2 , что приводит к образованию углеводов. Этот процесс фиксации СО 2 и образования углеводов состоит из многих этапов, в которых участвует большое число ферментов (цикл Кальвина). Биохимическими исследованиями было показано, что ферменты, участвующие в темновых реакциях, содержатся в водорастворимой фракции хлоропластов, содержащей компоненты матрикса-стромы этих пластид.
Процесс восстановления СО 2 начинается с его присоединения к рибулозодифосфату, углеводу, состоящему из 5 атомов углерода с образованием короткоживущего С 6 -соединения, которое сразу распадается на два С 3 -соединения, на две молекулы глицерид-3-фосфата.
Именно на этом этапе при карбоксилировании рибулозодифосфата и происходит связывание СО 2 . Дальнейшие реакции превращения глицерид-3-фосфата приводят к синтезу различных гексоз и пентоз, к регенерации рибулозодифосфата и к его новому вовлечению в цикл реакций связывания СО 2 . В конечном счете в хлоропласте из шести молекул СО 2 образуется одна молекула гексозы, для этого процесса требуется 12 молекул НАДФ-Н и 18 молекул АТФ, поступающих из световых реакций фотосинтеза. Образовавшийся в результате темновой реакции фруктоза-6 –фосфат дает начало сахарам, полисахаридам (крахмал) и галактолипидам. В строме хлоропластов кроме того из части глицерид-3-фосфата образуются жирные кислоты, аминокислоты и крахмал. Синтез сахарозы завершается в цитоплазме.
В строме хлоропластов происходит восстановление нитритов до аммиака, за счет энергии электронов, активированных светом; в растениях этот аммиак служит источником азота при синтезе аминокислот и нуклеотидов.
Онтогенез и функциональные перестройки пластид
Многих исследователей занимал вопрос о происхождении пластид и о путях их образования.
Еще в конце позапрошлого столетия было найдено, что у нитчатой зеленой водоросли спирогиры деление клеток при вегетативном размножении сопровождается делением их хроматофора путем перетяжки. Подробно исследована судьба хлоропласта у зеленой водоросли хламидомонады (рис. 230). Оказалось, что при бесполом, вегетативном размножении сразу вслед за делением ядра наступает перешнуровка гигантского хроматофора на две части, каждая из которых попадает в одну из дочерних клеток, где дорастает до исходной величины. Такое же равное разделение хлоропласта происходит и при формировании зооспор. При образовании зиготы после слияния гамет, каждая из которых содержала хлоропласт, после объединения ядер хлоропласты сначала соединяются тонкой перемычкой, а затем их содержимое сливается в одну крупную пластиду.
У высших растений также встречается деление зрелых хлоропластов, но очень редко. Увеличение числа хлоропластов и образование других форм пластид (лейкопластов и хромопластов) следует рассматривать как путь превращения структур-предшественников, пропластид . Весь же процесс развития различных пластид можно представить в виде монотропного (идущего в одном направлении) ряда смены форм:
Пропластида лейкопласт хлоропласт хромопласт
амилопласт
Многими исследованиями был установлен необратимый характер онтогенетических переходов пластид. У высших растений возникновение и развитие хлоропластов происходят через изменения пропластид (рис. 231).
Пропластиды представляют собой мелкие (0,4-1 мкм) двумембранные пузырьки, не имеющие отличительных черт их внутреннего строения. Они отличаются от вакуолей цитоплазмы более плотным содержимым и наличием двух отграничивающих мембран, внешней и внутренней (наподобие промитохондриям у дрожжевых клеток). Внутренняя мембрана может давать небольшие складки или образовывать мелкие вакуоли. Пропластиды чаще всего встречаются в делящихся тканях растений (клетки меристемы корня, листьев, в точки роста стеблей и др.). По всей вероятности, увеличение их числа происходит путем деления или почкования, отделения от тела пропластиды мелких двумембранных пузырьков.
Судьба таких пропластид будет зависеть от условий развития растений. При нормальном освещении пропластиды превращаются в хлоропласты . Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов.
Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид, этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов.
При освещении клеток мембранные пузырьки и трубочки быстро реорганизуются, из них развивается полная система ламелл и тилакоидов, характерная для нормального хлоропласта.
Лейкопласты отличаются от хлоропластов отсутствием развитой ламеллярной системы (рис. 226 б). Встречаются они в клетках запасающих тканей. Из-за их неопределенной морфологии лейкопласты трудно отличить от пропластид, а иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но тем не менее способны к образованию под влиянием света нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать в проламеллярных телах различные запасные вещества, а в строме лейкопластов откладываются зерна вторичного крахмала. Если в хлоропластах происходит отложение так называемого транзиторного крахмала, который присутствует здесь лишь во время ассимиляции СО 2 , то в лейкопластах может происходить истинное запасание крахмала. В некоторых тканях (эндосперм злаков, корневища и клубни) накопление крахмала в лейкопластах приводит к образованию амилопластов , сплошь заполненных гранулами запасного крахмала, расположенных в строме пластиды (рис. 226в).
Другой формой пластид у высших растений является хромопласт, окрашивающийся обычно в желтый свет в результате накопления в нем каротиноидов (рис. 226г). Хромопласты образуются из хлоропластов и значительно реже их лейкопластов (например, в корне моркови). Процесс обесцвечивания и изменения хлоропластов легко наблюдать при развитии лепестков или при созревании плодов. При этом в пластидах могут накапливаться окрашенные в желтый цвет капельки (глобулы) или в них появляются тела в форме кристаллов. Эти процессы сопряжены с постепенным уменьшением числа мембран в пластиде, с исчезновением хлорофилла и крахмала. Процесс образования окрашенных глобул объясняется тем, что при разрушении ламелл хлоропластов выделяются липидные капли, в которых хорошо растворяются различные пигменты (например, каротиноиды). Таким образом, хромопласты представляют собой дегенерирующие формы пластид, подвернутые липофанерозу – распаду липопротедных комплексов.
Фотосинтезирующие структуры низших эукариотических и
прокариотических клеток
Строение пластид у низших фотосинтезирующих растений (зеленые, бурые и красные водоросли) в общих чертах сходно с хлоропластами клеток высших растений. Их мембранные системы также содержат фоточувствительные пигменты. Хлоропласты зеленых и бурых водорослей (иногда их называют хроматофорами) имеют также внешнюю и внутреннюю мембраны; последняя образует плоские мешки, располагающиеся параллельными слоями, граны у этих форм не встречаются (рис. 232). У зеленых водорослей в состав хроматофора входят пиреноиды , представляющие собой окруженные мелкими вакуолями зону, вокруг которой происходит отложение крахмала (рис. 233).
Форма хлоропластов у зеленых водорослей очень разнообразна – это или длинные спиральные ленты (Spirogira), сети (Oedogonium), или мелкие округлые, похожие на хлоропласты высших растений (рис. 234).
Среди прокариотических организмов многие группы обладают фотосинтетическими аппаратами и имеют в связи с этим особое строение. Для фотосинтезирующих микроорганизмов (синезеленые водоросли и многие бактерии) характерно, что их фоточувствительные пигменты локализуются в плазматической мембране или в её выростах, направленных вглубь клетки.
В мембранах синезеленых водорослей кроме хлорофилла находятся пигменты фикобилины. Фотосинтезирующие мембраны синезеленых водорослей образуют плоские мешки (ламеллы), которые располагаются параллельно друг над другом, иногда образуя стопки или спирали. Все эти мембранные структуры образуются за счет инвагинаций плазматической мембраны.
У фотосинтезирующих бактерий (Chromatium) мембраны образуют мелкие пузырьки, число которых так велико, что они заполняют практически большую часть цитоплазмы. Можно видеть, как эти пузырьки образуются за счет впячивания и последующего роста плазматической мембраны. Эти мембранные пузырьки (их также называют хроматофорами) содержат фоточувствительный пигмент бактериохлорофилл, каратиноиды, компоненты фотосинтетической системы переноса электронов и фотофосфорилирования. Некоторые пурпурные бактерии содержат систему мембран, образующих правильные стопки, наподобие тилакоидов в гранах хлоропластов (рис. 235).
Геном пластид
Подобно митохондриям, хлоропласты имеют собственную генетическую систему, обеспечивающую синтез ряда белков внутри самих пластид. В матриксе хлоропластов обнаруживаются ДНК, разные РНК и рибосомы. Оказалось, что ДНК хлоропластов резко отличается от ДНК ядра. Она представлена циклическими молекулами длиной до 40-60 мкм, имеющими молекулярный вес 0,8-1,3х10 8 дальтон. В одном хлоропласте может быть множество копий ДНК. Так, в индивидуальном хлоропласте кукурузы присутствует 20-40 копий молекул ДНК. Длительность цикла и скорость репликации ядерной и хлоропластной ДНК, как было показано на клетках зеленых водорослей, не совпадают. ДНК хлоропластов не состоит в комплексе с гистонами. Все эти характеристики ДНК хлоропластов близки к характеристикам ДНК прокариотических клеток. Более того, сходство ДНК хлоропластов и бактерий подкрепляется еще и тем, что основные регуляторные последовательности транскрипции (промоторы, терминаторы) у них одинаковы. На ДНК хлоропластов синтезируются все виды РНК (информационная, трансферная, рибосомная). ДНК хлоропластов кодирует рРНК, входящую в состав рибосом этих пластид, которые относятся к прокариотическому 70S типу (содержат 16S и 23S рРНК). Рибосомы хлоропластов чувствительны к антибиотику хлорамфениколу, подавляющему синтез белка у прокариотических клеток.
Так же как в случае хлоропластов мы вновь сталкиваемся с существованием особой системы синтеза белка, отличной от таковой в клетке.
Эти открытия вновь пробудили интерес к теории симбиотического происхождения хлоропластов. Идея о том, что хлоропласты возникли за счет объединения клеток-гетеротрофов с прокариотическими синезелеными водорослями, высказанная на рубеже XIX и XX вв. (А.С. Фоминцин, К.С.Мережковский) вновь находит свое подтверждение. В пользу этой теории говорит удивительное сходство в строении хлоропластов и синезеленых водорослей, сходство с основными их функциональными особенностями, и в первую очередь со способностью к фотосинтетическим процессам.
Известны многочисленные факты истинного эндосимбиоза синезеленых водорослей с клетками низших растений и простейших, где они функционируют и снабжают клетку-хозяина продуктами фотосинтеза. Оказалось, что выделенные хлоропласты могут также отбираться некоторыми клетками и использоваться ими как эндосимбионты. У многих беспозвоночных (коловратки, моллюски), питающихся высшими водорослями, которые они переваривают, интактные хлоропласты оказываются внутри клеток пищеварительных желез. Так, у некоторых растительноядных моллюсков в клетках найдены интактные хлоропласты с функционирующими фотосинтетическими системами, за активностью которых следили по включению С 14 О 2 .
Как оказалось, хлоропласты могут быть введены в цитоплазму клеток культуры фибробластов мыши путем пиноцитоза. Однако они не подвергались атаке гидролаз. Такие клетки, включившие зеленые хлоропласты, могли делиться в течение пяти генераций, а хлоропласты при этом оставались интактными и проводили фотосинтетические реакции. Были предприняты попытки культивировать хлоропласты в искусственных средах: хлоропласты могли фотосинтезировать, в них шел синтез РНК, они оставались интактными 100 ч, у них даже в течение 24 ч наблюдались деления. Но затем происходило падение активности хлоропластов, и они погибали.
Эти наблюдения и целый ряд биохимических работ показали, что те черты автономии, которыми обладают хлоропласты, еще недостаточны для длительного поддержания их функций и тем более для их воспроизведения.
В последнее время удалось полностью расшифровать всю последовательность нуклеотидов в составе циклической молекулы ДНК хлоропластов высших растений. Эта ДНК может кодировать до 120 генов, среди них: гены 4 рибосомных РНК, 20 рибосомных белков хлоропластов, гены некоторых субъединиц РНК-полимеразы хлоропластов, несколько белков I и II фотосистем, 9 из 12 субъединиц АТФ-синтетазы, части белков комплексов цепи переноса электронов, одной из субъединиц рибулозодифосфат-карбоксилазы (ключевой фермент связывания СО 2), 30 молекул тРНК и еще 40 пока неизвестных белков. Интересно, что сходный набор генов в ДНК хлоропластов обнаружен у таких далеко отстоящих представителей высших растений как табак и печеночный мох.
Основная же масса белков хлоропластов контролируется ядерным геномом. Оказалось, что ряд важнейших белков, ферментов, а соответственно и метаболические процессы хлоропластов находятся под генетическим контролем ядра. Так, клеточное ядро контролирует отдельные этапы синтеза хлорофилла, каротиноидов, липидов, крахмала. Под ядерным контролем находятся многие энзимы темновой стадии фотосинтеза и другие ферменты, в том числе некоторые компоненты цепи транспорта электронов. Ядерные гены кодируют ДНК-полимеразу и аминоацил-тРНК-синтетазу хлоропластов. Под контролем ядерных генов находится большая часть рибосомных белков. Все эти данные заставляют говорить о хлоропластах, так же как и о митохондриях, как о структурах с ограниченной автономией.
Транспорт белков из цитоплазмы в пластиды происходит в принципе сходно с таковым у митохондрий. Здесь также в местах сближения внешней и внутренней мембран хлоропласта располагаются каналообразующие интегральные белки, которые узнают сигнальные последовательности хлоропластных белков, синтезированных в цитоплазме, и транспортируют их в матрикс-строму. Из стромы импортируемые белки согласно дополнительным сигнальным последовательностям могут включаться в мембраны пластиды (тилакоиды, ламеллы стромы, внешняя и внутренняя мембраны) или локализоваться в строме, входя в состав рибосом, ферментных комплексов цикла Кальвина и др.
Удивительное сходство структуры и энергетических процессов у бактерий и митохондрий, с одной стороны, и у синезеленых водорослей и хлоропластов – с другой, служит веским аргументом в пользу теории симбиотического происхождения этих органелл. Согласно этой теории, возникновение эукариотической клетки прошло через несколько этапов симбиоза с другими клетками. На первой стадии клетки типа анаэробных гетеротрофных бактерий включили в себя аэробные бактерии, превратившиеся в митохондрии. Параллельно этому в клетке-хозяине прокариотический генофор формируется в обособленное от цитоплазмы ядро. Так могли возникнуть гетеротрофные эукариотические клетки. Повторные эндосимбиотические взаимоотношения между первичными эукариотическими клетками и синезелеными водорослями привели к появлению в них структур типа хлоропластов, позволяющих клеткам осуществлять автосинтетические процессы и не зависеть от наличия органических субстратов (рис. 236). В процессе становления такой составной живой системы часть генетической информации митохондрий и пластид могла изменяться, перенестись в ядро. Так, например две трети из 60 рибосомных белков хлоропластов кодируется в ядре и синтезируются в цитоплазме, а потом встраивается в рибосомы хлоропластов, имеющие все свойства прокариотических рибосом. Такое перемещение большой части прокариотических генов в ядро привело к тому, что эти клеточные органеллы, сохранив часть былой автономии, попали под контроль клеточного ядра, определяющего в большей степени все главные клеточные функции.